Desolee d'ecrire en anglais, but my french is so bad, so I hope you excuse me to write in English, if not in japanese.
Salut, Mon Ami
I don't know how many people here are really interested in who the
japanese people really are. But since this is the forum about japan,
I think it nice idea to post some recent
scientific study concerning japanese people.
The stated conclustion from many various studies is:
(1) 60-70% of Japanese population are actually from Korea
(2) 30-40% of Japanese population are native islanders from northern sibelia
I am planning to post some genetic studies to show that the statement above is true.
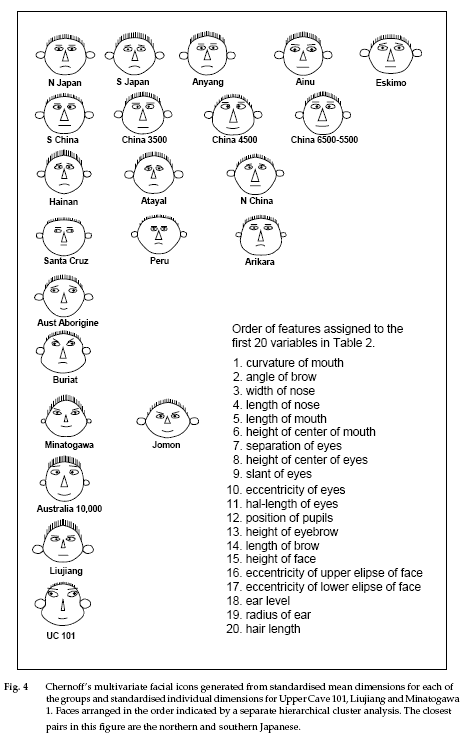
I will then talk about the facial structure of Japanese people in relation to Korean and Chinese. Lastly, I will post some pictures of Japanese People so that you could see the morphological differences between the groups.
-------------------
: J Hum Genet. 2000;45(2):76-83. Related Articles, Links
Y chromosomal DNA variation in east Asian populations and its potential for inferring the peopling of Korea.
Kim W, Shin DJ, Harihara S, Kim YJ.
Department of Biology, Dankook University, Cheonan, Choong-Nam, Republic of Korea. wookkim@ansco.dankook.ac.kr
We have examined variations of five polymorphic loci (DYS287, DXYS5Y, SRY465, DYS19, and DXYS156Y) on the Y chromosome in samples from a total of 1260 males in eight ethnic groups of East Asia. We found four unique haplotypes constructed from three biallelic markers in these samples of East Asians. The Japanese population was characterized by a relatively high frequency of either the haplotype I-2b (-/Y2/T) or II-1 (+/Y1/C). These dual patterns of the distribution of Y chromosomes (I-2b/II-1) were also found in Korea, although they were present at relatively low frequencies. The haplotype II-1 was present in Northeast Asian populations (Chinese, Japanese, Koreans, and Mongolians) only, except for one male from the Thai population among the Southeast Asian populations (Indonesians, Philippines, Thais, and Vietnamese). The Japanese were revealed to have the highest frequency of this haplotype (27.5%), followed by Koreans (2.9%), Mongolians (2.6%), and mainland Chinese (2.2%). In contrast, the frequency of the haplotype I-2b was found to be 17.1% in the Japanese, 9.5% in Indonesian, 6.3% in Korean, 3.8% in Vietnamese, and 2.7% in Thai samples. These findings suggested that the chromosomes of haplotype I-2b were likely derived from certain areas of Northeast Asia, the region closest to Southeast Asia. Phylogenetic analysis using the neighbor-joining tree also reflected a general distinction between Southeast and Northeast Asian populations. The phylogeny revealed a closer genetic relationship between Japanese and Koreans than to the other surveyed Asian populations. Based on the result of the dual patterns of the haplotype distribution, it is more likely that the population structure of Koreans may not have evolved from a single ancient population derived from Northeast Asians, but through dual infusions of Y chromosomes entering Korea from two different waves of East Asians.
PMID: 10721667 [PubMed - indexed for MEDLINE]
Fig. 2 Distribution of Y haplogroups in east Asia. Circle area is proportional to sample size, and the nine haplogroups are represented by different colors
The distribution of Y-chromosomal variation surveyed here reveals significant genetic differences among east Asian populations. Haplogroup DE-YAP (the YAP+ allele) was present at high frequency only in the Japanese and was rare in other parts of east Asia (Table 2, Fig. 2). This result is consistent with previous findings of YAP+ chromosomes only in populations from Japan and Tibet in east Asia (Hammer and Horai 1995; Hammer et al. 1997; Kim et al. 2000; Tajima at al. 2002). However, haplogroup DE-YAP is also found at low frequencies in all the other northeast Asian populations sampled here (2.4% overall, excluding the Japanese; 9.6%, including the Japanese), but only in two of the southern populations (0.8% overall), suggesting that the Korean YAP+ chromosomes are unlikely to have been derived from a southeast Asian source. The prevalence of the YAP+ allele in central Asian populations suggests a genetic contribution to the east Asian populations from the northwest, probably from central Asia (Altheide and Hammer 1997; Jin and Su 2000; Karafet et al. 2001).
Haplogroups C-RPS4Y711 and K-M9 were widely but not evenly distributed in the east Asian populations. Haplogroup C-RPS4Y711 appears to be the predominant northeast Asian haplogroup, with high frequencies in Mongolians (Buryats, 37.3%; Khalkhs, 42.9%) and Manchurians (22.7%; Table 2, Fig. 2). The moderate frequency of haplogroup C-RPS4Y711 Y-chromosomes in Korea (15.0%) implies a genetic influence from northern populations of east Asia, starting possibly in east Siberia. Su and Jin (2001) suggest that the RPS4Y711-T chromosome originated in east Asia, probably in the southeast, and then expanded to the north (Siberia), based on the genetic diversity of Y-STR markers. However, the observed low Y-STR diversity of haplogroup C-RPS4Y711 chromosomes in their surveys of Siberian and central Asian populations compared with east Asian populations could also be explained by a more northern (Mongolian and/or Siberian) origin followed by genetic drift resulting from small effective population sizes (Pakendorf et al. 2002). Recently, Cavalli-Sforza and Feldman (2003) have suggested that haplogroup C-RPS4Y711 expanded both through a southern route from Africa (e.g., India) to Oceania, and a northern one to Mongolia, Siberia, and eventually to northwest America. Further genetic surveys are required to test these hypotheses, with additional markers and more samples from diverse regions of Asia.
In contrast, M9-G Y-chromosomes show an opposing distribution to those carrying RPS4Y711-T in east Asia: they are more frequent in southern populations than in northern ones, showing a clinal variation from about 90% to 60% (Table 1). The haplogroups carrying the M9-G mutation and additional sublineages of M9-G in Korea appear to be at an intermediate frequency (81.9%) between southeast and northeast Asian populations. This result implies that the Korean population may be influenced by both the northeast and southeast Asian populations. Even within haplogroup O, the most frequent Korean STR haplotype (23-10-13 with the markers DYS390-DYS391-DYS393, 19% of haplogroup O; Table 3) is the most frequent in the Philippines (27%), whereas the second most frequent Korean haplotype (24-10-12, 16%) is the most frequent in Manchuria (45%). Thus, the distribution of haplogroups K-M9 and C-RPS4Y711 may reflect dispersals from both north and south. The settlement of each region at different times needs to be considered in order to understand the peopling of east Asia. Recently, Karafet et al. (2001) have noted that realistic explanations for the peopling of east Asia have to accommodate more complex multidirectional biological and cultural influences than earlier models have allowed.
Fig. 3 Principal components (PC) analysis of haplogroup frequencies in 11 east Asian populations (circle Koreans, open diamonds southeast populations, closed diamonds northeast populations)
In this study, the Koreans appear to be most closely related overall to the Manchurians among east Asian ethnic groups (Fig. 2), although a principal components analysis of haplogroup frequencies reveals that they also cluster with populations from Yunnan and Vietnam (Fig. 3). The genetic relationship with Manchuria is consistent with the historical evidence that the Ancient Chosun, the first state-level society, was established in the region of southern Manchuria and later moved into the Pyongyang area of the northwestern Korean Peninsula. Based on archeological and anthropological data, the early Korean population possibly had a common origin in the northern regions of the Altai Mountains and Lake Baikal of southeastern Siberia (Han 1995; Choi and Rhee 2001). Recent studies of mtDNA (Kivisild et al. 2002) and the Y-chromosome (Karafet et al. 2001) have also indicated that Koreans possess lineages from both the southern and the northern haplogroup complex. In conclusion, the peopling of Korea can be seen as a complex process with an initial northern Asian settlement followed by several migrations, mostly from southern-to-northern China.[/img]